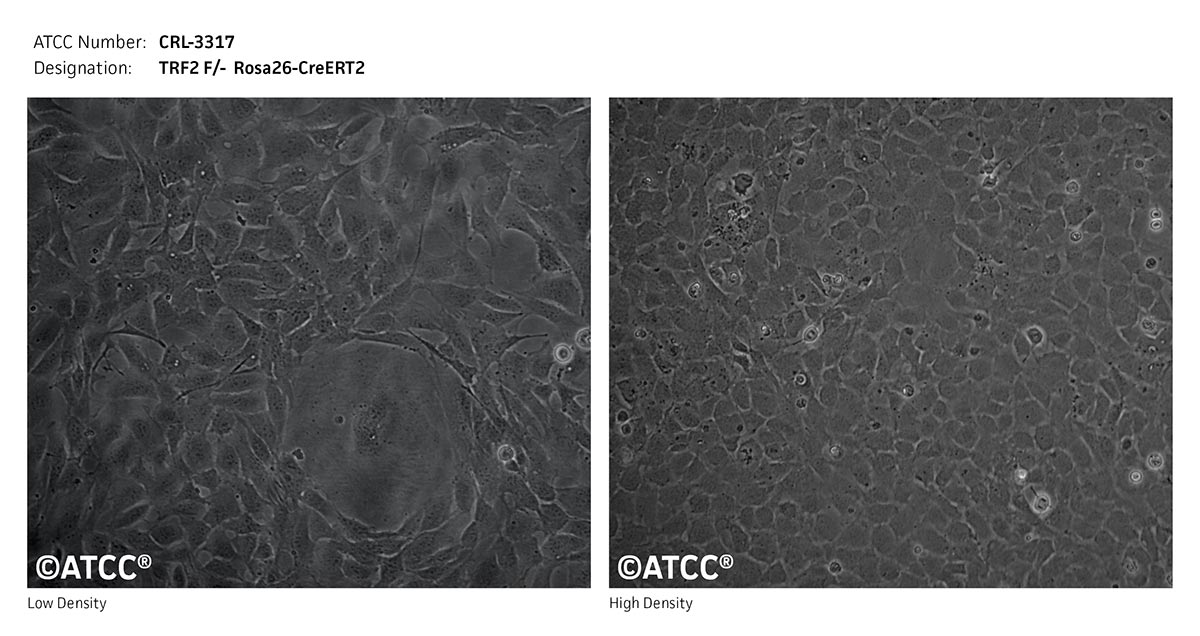
TRF2 F/- Rosa26-CreERT2
CRL-3317 ™
CRL-3317 ™
ATCC determines the biosafety level of a material based on our risk assessment as guided by the current edition of Biosafety in Microbiological and Biomedical Laboratories (BMBL), U.S. Department of Health and Human Services. It is your responsibility to understand the hazards associated with the material per your organization’s policies and procedures as well as any other applicable regulations as enforced by your local or national agencies.
Cells contain retroviral sequences
Cells contain SV40 sequences
ATCC highly recommends that appropriate personal protective equipment is always used when handling vials. For cultures that require storage in liquid nitrogen, it is important to note that some vials may leak when submersed in liquid nitrogen and will slowly fill with liquid nitrogen. Upon thawing, the conversion of the liquid nitrogen back to its gas phase may result in the vial exploding or blowing off its cap with dangerous force creating flying debris. Unless necessary, ATCC recommends that these cultures be stored in the vapor phase of liquid nitrogen rather than submersed in liquid nitrogen.
This mouse embryonic fibroblast cell line possess loxP sites on either side of the first and second coding exon of mouse TRF2 (Terf2) gene. This MEF line also expresses Cre-ERT2 from one Rosa26 locus. TRF2 knockout can be done by Cre recombinase transduction or 0.5 uM 4-hydroxytamoxifen treatment for 6hr of the MEFs. This inducible TRF2 knockout MEF line is useful in studies of telomere biology.
Development of the system:
A targeting vector containing neomycin resistance and herpes simplex virus thymidine kinase genes under the control of the phosphoglyceryl kinase gene promoter was utilized in the construction of this mutant. A loxP site was inserted upstream of exon 1. Another loxP site was inserted downstream of exon 2. This construct was electroporated into 129P2/OlaHsd derived E14 embryonic stem (ES) cells which were transiently transfected with a Cre recombinase vector to remove the selection cassette. Correctly targeted ES cells were injected into recipient blastocysts.
To ensure the highest level of viability, thaw the vial and initiate the culture as soon as possible upon receipt. If upon arrival, continued storage of the frozen culture is necessary, it should be stored in liquid nitrogen vapor phase and not at -70°C. Storage at -70°C will result in loss of viability.
The product is provided 'AS IS' and the viability of ATCC® products is warranted for 30 days from the date of shipment, provided that the customer has stored and handled the product according to the information included on the product information sheet, website, and Certificate of Analysis. For living cultures, ATCC lists the media formulation and reagents that have been found to be effective for the product. While other unspecified media and reagents may also produce satisfactory results, a change in the ATCC and/or depositor-recommended protocols may affect the recovery, growth, and/or function of the product. If an alternative medium formulation or reagent is used, the ATCC warranty for viability is no longer valid. Except as expressly set forth herein, no other warranties of any kind are provided, express or implied, including, but not limited to, any implied warranties of merchantability, fitness for a particular purpose, manufacture according to cGMP standards, typicality, safety, accuracy, and/or noninfringement.
This product is intended for laboratory research use only. It is not intended for any animal or human therapeutic use, any human or animal consumption, or any diagnostic use. Any proposed commercial use is prohibited without a license from ATCC.
While ATCC uses reasonable efforts to include accurate and up-to-date information on this product sheet, ATCC makes no warranties or representations as to its accuracy. Citations from scientific literature and patents are provided for informational purposes only. ATCC does not warrant that such information has been confirmed to be accurate or complete and the customer bears the sole responsibility of confirming the accuracy and completeness of any such information.
This product is sent on the condition that the customer is responsible for and assumes all risk and responsibility in connection with the receipt, handling, storage, disposal, and use of the ATCC product including without limitation taking all appropriate safety and handling precautions to minimize health or environmental risk. As a condition of receiving the material, the customer agrees that any activity undertaken with the ATCC product and any progeny or modifications will be conducted in compliance with all applicable laws, regulations, and guidelines. This product is provided 'AS IS' with no representations or warranties whatsoever except as expressly set forth herein and in no event shall ATCC, its parents, subsidiaries, directors, officers, agents, employees, assigns, successors, and affiliates be liable for indirect, special, incidental, or consequential damages of any kind in connection with or arising out of the customer's use of the product. While reasonable effort is made to ensure authenticity and reliability of materials on deposit, ATCC is not liable for damages arising from the misidentification or misrepresentation of such materials.
Please see the material transfer agreement (MTA) for further details regarding the use of this product. The MTA is available at www.atcc.org.
For-profit Organizations: For every order of this item, you must work directly with the contributor, Rockefeller University to (i) negotiate a research-use license, and/or (ii) have the contributor provide authorization to ATCC to ship this item under your existing license. We cannot ship this item until we receive communication directly from Rockefeller University that we are authorized to ship each order.
We are providing the following contact information, but this information may change without notice:
Rockefeller University
Office of Technology Transfer
Attn. Nidhi Sabharwal, Assistant Director, Marketing & Licensing
1230 York Avenue, New York, NY 10065
Email: [email protected]
Once ATCC has received authorization from Rockefeller University, your order will be reviewed, and this item will be released for shipment if all requirements are met. If you need assistance with your order, please contact our Customer Care team or your applicable distributor.
If shipping to the U.S. state of Hawaii, you must provide either an import permit or documentation stating that an import permit is not required. We cannot ship this item until we receive this documentation. Contact the Hawaii Department of Agriculture (HDOA), Plant Industry Division, Plant Quarantine Branch to determine if an import permit is required.
Dimitrova N, et al. 53BP1 promotes non-homologous end joining of telomeres by increasing chromatin mobility. Nature 456(7221): 524-528, 2008. PubMed: 18931659
Celli G, de Lange T. DNA processing not required for ATM activation or the telomere damage response after conditional deletion of mouse TRF2. Nat Cell Biol 7(7): 712-718, 2005. PubMed: 15968270

 Powered by Bioz
Powered by Bioz
